

Research Article
Received: 2026-02-19 | Revised:2026-04-16 | Accepted: 2026-04-19 | Published: 2026-05-25
Pages: 52-62
DOI: https://doi.org/10.58985/jpam.2026.v04i01.44
Abstract
Kribi is one of Cameroon's most popular seaside resorts. Its beaches are generally fed by rivers and are sites of numerous human activities. Aquatic environments are increasingly subject to multiple forms of human-induced contamination. The pathogenic microorganisms present can cause illness and economic losses in activities that depend primarily on seawater. This study aimed to determine the susceptibility of two species of the genus Staphylococcus and the potential impact of proximity/contact with seawater on the antibiotic susceptibility of these two Staphylococcus species isolated from underground and surface coastal waters at Kribi (Cameroon). The bacteria were isolated using the surface spread technique on Petri dishes with Chapman mannitol agar (HIMEDIA, ref. MH118-500G). The isolated bacteria were identified using standard methods. The susceptibility of the bacteria to antibiotics was verified using the disk diffusion method on Müller-Hinton agar, following the recommendations of the Antibiogram Committee of the French Society of Microbiology. Bacteriological analyses revealed that these waters host a bacterial microflora composed of Staphylococcus aureus and Staphylococcus epidermidis. Antibiotic susceptibility tests showed that the different bacterial species exhibited resistance to methicillin (100%) (due to their resistance to cefoxitin) and penicillin G, which belongs to the β-lactam family. Within the quinolone, aminoglycoside, phenicol, and diaminopyrimidine/sulfamide families, the percentages of antibiotic susceptibility for the different species of Staphylococcus aureus and Staphylococcus epidermidis were all greater than 50% on average, regardless of the biotope. The percentage of staphylococcal resistance in underground and coastal waters was higher (78.57%), compared to surface waters (71.43%). The susceptibility to gentamicin in the different isolated staphylococcal species appeared to be favored by their proximity or contact with seawater, thus showing the influence of abiotic factors on the antibiotic susceptibility of bacterial species.
Keywords
Surface water, underground water, coastal water, Staphylococcus, antibiotic.
1. Introduction
Staphylococci
are commensal bacteria found on the skin and mucous membranes of humans and
animals [1]. Humans are the primary natural
reservoir for Staphylococcus, and
they are healthy carriers, mainly in the nasal cavity. In humans,
staphylococci, particularly Staphylococcus
aureus and Staphylococcus epidermidis,
are part of the resident cutaneous flora of many individuals who are
"asymptomatic carriers" [2]. Infections
caused by Staphylococcus aureus are
known to be responsible for numerous skin conditions (abscesses, boils, and
paronychia) or mucous membrane infections (otitis and conjunctivitis), which
are more severe and therefore more difficult to treat [3].
The presence of staphylococci in water can be due to the contamination of the
aquatic environment by fecal pollution or by contamination occurring during the
processing or transformation of seafood [4].
Human contamination occurs through drinking water consumption, consumption of
food contaminated by water (fish, seafood, etc.), or during bathing or contact
with recreational water [5]. Numerous
studies have been conducted on the microbiological quality of beach waters in
Cameroon, particularly in the seaside towns of Kribi and Limbé. Indeed, these
analyses have revealed the presence of fecal pollution indicator germs and
other pathogenic microorganisms, such as Aeromonas
sp., Bacillus sp., Pseudomonas sp., Staphylococcus sp., and Vibrio
sp. Infections caused by resistant bacterial species are generally more
severe and more difficult to treat [3]. Numerous
studies have been conducted on the microbiological quality of beach water in
Cameroon, particularly in the seaside towns of Kribi and Limbé. Indeed, these
analyses have revealed the presence of fecal pollution indicator germs and
other pathogenic microorganisms such as Aeromonas
sp., Bacillus sp., Pseudomonas sp., Staphylococcus sp., and Vibrio
sp. in these waters. The abundance dynamics of these microorganisms vary
seasonally, and these waters are not recommended and therefore of poor quality
for swimming [5]. Similarly, information on
the comparative abundance of Staphylococcus
species in three different aquatic systems in Kribi has been published. These
studies indicate that these waters, which are neutral and poorly mineralized,
host a bacterial microflora composed of Staphylococcus
aureus and Staphylococcus epidermidis
as the high proportions [6]. The work
carried out by Tamsa Arfao et al. [7] in the
waters of some wastewater treatment plants in Yaoundé-Cameroon revealed the
presence of bacteria of the genus Staphylococcus
resistant to many antibiotics. Previously, it showed that
waterborne diseases cause gastroenteritis, skin and eye infections in swimmers,
urinary tract infections, and meningitis, among others [5,
8-10]. These diseases are responsible for a high mortality rate in emerging
countries [11]. Given the
presence of these germs in these waters, the most appropriate antibiotics were sulfamethoxazole/trimethoprim
(35%) and ciprofloxacin (32.5%), while the most resistant were ampicillin
(57.5%), followed by ofloxacin (55%) and amoxicillin (50%) [12-13]. This information sheds light on the
issues concerning the updating of data on the antibiotic susceptibility of
bacteria present in coastal and continental waters used as alternatives by
populations in emerging countries. Similarly, there are few updated data on the
antibiotic susceptibility of Staphylococcus
species present in the coastal and continental waters of Kribi, which are
regularly used by the resident population when accessible. Furthermore, the
potential impact of proximity/contact with seawater on the susceptibility of
strains isolated from continental waters has been little studied. This study
aimed to determine the impact of proximity/contact with seawater on the
antibiogram susceptibility of two Staphylococcus
species isolated from underground, surface and coastal waters in Kribi
(Cameroon).
2. Materials and methods
Eight sampling stations divided into three aquatic systems (underground surface and coastal waters), were chosen. For undergrounds water, two boreholes coded (B1 and B2) and two wells (W1 and W2) were selected. For surfaces two points (coded N1 and N2) located in the Nlendé river were chosen. For seawater, two points (coded S1 to S2) were also selected. Fig. 1 showed a map of the sampling site locations. The geographical coordinates of each sampling point are listed in Table 1. It was observed that near these sampling points, there were punctual and continuous pollution sources originating from toilet discharges and anthropogenic activities carried out by port structures located not far from the studied river.

Figure 1: Location of
the different sampling points in the city of Kribi.
Table 1. Codes, locations and distances from the sea of the different sampling points.
|
Codes |
B1 |
B2 |
W1 |
W2 |
N1 |
N2 |
S1 |
S2 |
|
Localisation |
Neighbor-hood
of Nziou |
Eboudja
village |
Neighbor-hood
of Bikondo |
Neighbor-hood
of Ngoyé |
Nlendé
village |
Nlendé
village |
Neighbor-hood of Ngoyé |
Port
of Mboro |
|
Distance from
the sea (m) |
128 |
214 |
201 |
81 |
296 |
16 |
0 |
0 |
|
The samples were taken in March 2024 in sterile 500 mL glass bottles for bacteriological analysis. The different samples were collected according to the techniques recommended by APHA [14] and Rodier [15]. |
||||||||
2.1. Evaluation of the susceptibility of isolated staphylococci to antibiotics
Isolation was performed using the membrane filtration technique for underground water and surface spread for surface and seawater samples on selective Chapman mannitol agar. Identification of presumptive strains was carried out using classical biochemical gallery methods [16].
The choice of antibiotics tested on Staphylococcus species was based on the recommendations of the CA-SFM/EUCAST [17]. A total of 7 distinct antibiotics were selected for this purpose, belonging to: the β-lactam, quinolone and aminoglycoside, sulfamides and other families. These antibiotics were chosen based on clinical indications and prevalence of acquired resistance [17]. The medium used was Müller-Hinton agar, which was sterilized by autoclaving at 121°C for 20 min, then poured into Petri dishes. The agar depth was approximately 4 mm [17]. The bacterial suspension was prepared from an 18–24 h culture on non-selective agar medium (plate count agar) and a saline solution (0.9% NaCl). The turbidity was adjusted to 1 x 10⁸ CFU/mL by spectrophotometry at a wavelength of 600 nm. This turbidity corresponds to that of the MacFarland standard 0.5, with an optical density between 0.08 and 0.1. The inoculum was then diluted 1/100 (1 x 10⁶ CFU/mL) before being seeded onto Müller-Hinton agar [17].
The agar was inoculated with the bacterial inoculum by the swabbing method. The entire surface of the agar was swabbed in three directions, then the Petri dishes were placed under a hood for 10 min. After drying, the antibiotic disks were placed on the surface of the agar with a 3 cm spacing to avoid overlapping of inhibition zones. The Petri dishes were left at room temperature for pre-diffusion of the antibiotics, then incubated for 10 min after, at 37°C for 18 to 24 h [17].
The diameters of the circular inhibition zones were measured to the nearest millimeter using a caliper. The interpretation of these diameters was based on the breakpoint diameters provided by CA-SFM [18]. Strains were clinically categorized as resistant (R), sensitive (S), or intermediate sensitivity (I).
The percentage (%) of each clinical category was determined relative to the total number of strains tested. These percentages were calculated for each antibiotic using the following formula:
Resistance (%) = xr/t × 100; % Intermediate sensitivity = xi/t × 100
Sensitivity (%) = xs/t × 100
xr: sum of strains resistant to the antibiotic; xi: sum of strains showing intermediate sensitivity to the antibiotic; xs: sum of strains sensitive to the antibiotic; t: sum of strains tested for the antibiotic [18].
2.2. Data analyses
To verify the impact of the proximity of the different sampling points to the sea on the susceptibility of bacteria from the waters of the studied coastal area, Spearman's rank correlation coefficient was determined using SPSS 20.0 software. Comparisons between the considered variables were performed using Kruskal-Wallis "H" tests and Mann-Whitney "U" tests using PAST 2011 software.
3. Results
3.1. Susceptibility of Staphylococcus to antibiotics
3.1.1. Identified species
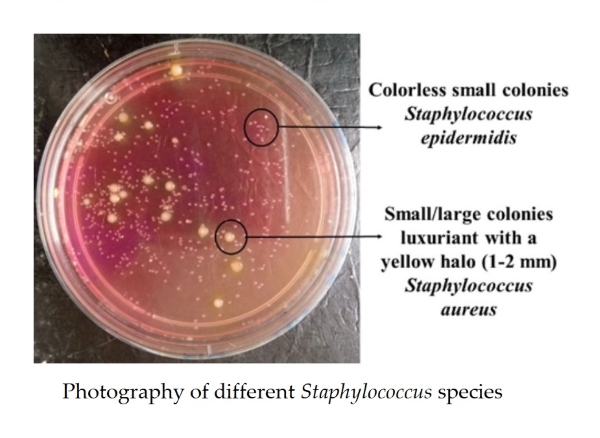
Examination of bacterial colonies across the all water points along the Kribi coast, revealed two types of colonies. Small/large colonies (diameter between 1 and 2 mm), luxuriant with a yellow halo, spherical in shape, capable of fermenting mannitol, reducing glucose and lactose, and coagulating rabbit plasma, correspond to Staphylococcus aureus. Colorless small colonies with almost the same biochemical characteristics as Staphylococcus aureus were those of Staphylococcus epidermidis (Fig. 2).

Figure 2. Photography of different Staphylococcus species.
3.1.2. Susceptibility of staphylococci to antibiotics
Antibiotic susceptibility was tested using different bacterial strains. The inhibition diameters for each isolated staphylococcal species from the different coastal, underground, and surface water samples are presented in Table 2. Overall, the inhibition zone diameters obtained ranged from 8‒33 mm for Staphylococcus aureus; and from 9‒30 mm for Staphylococcus epidermidis. In groundwater, for Staphylococcus aureus, the highest inhibition value (30 mm) was obtained with gentamicin at stations B2 and W2, whereas the lowest inhibition value (8 mm) was obtained with trimethoprim-sulfamethoxazole, at station W2. In contrast, for Staphylococcus epidermidis, the highest inhibition value (30 mm) was obtained with gentamicin and chloramphenicol at station B2, as well as with ciprofloxacin and norfloxacin at station W2. The lowest inhibition value (9 mm) was also obtained with trimethoprim-sulfamethoxazole at station W2. In surface waters, the highest inhibition value (32 mm) for Staphylococcus aureus was obtained with chloramphenicol at station N1, and the lowest value (15 mm) was obtained with penicillin G, also at station N1. For Staphylococcus epidermidis, the highest (30 mm) and lowest (12 mm) inhibition values were obtained with trimethoprim-sulfamethoxazole at stations N2 and N1, respectively. In coastal waters, the highest inhibition value (33 mm) was obtained with ciprofloxacin and chloramphenicol at station S2 for Staphylococcus aureus, and the lowest inhibition value (13 mm) was obtained with penicillin G at station S1. In Staphylococcus epidermidis, the highest inhibition value (25 mm) was obtained only with ciprofloxacin at station S2, and the lowest value (11 mm) was obtained with trimethoprim-sulfamethoxazole at station S2.
Table 2. Inhibition diameter (mm) of the different isolated Staphylococcus species according to the type of biotope.
|
Biotope
types |
Sampling points |
Species |
Inhibition zone (diameter in mm) |
||||||
|
FOX30 |
P10 |
CIP 5 |
NOR10 |
CN 10 |
SXT 25 |
C30 |
|||
|
Groundwater |
B1 |
S. aureus |
18 |
16 |
23 |
21 |
22 |
16 |
26 |
|
S. epidermidis |
16 |
20 |
23 |
22 |
23 |
14 |
20 |
||
|
B2 |
S. aureus |
17 |
13 |
24 |
24 |
30 |
16 |
29 |
|
|
S. epidermidis |
19 |
15 |
24 |
23 |
28 |
13 |
28 |
||
|
W1 |
S. aureus |
16 |
13 |
20 |
18 |
24 |
16 |
23 |
|
|
S. epidermidis |
22 |
14 |
20 |
17 |
23 |
12 |
23 |
||
|
W2 |
S. aureus |
20 |
16 |
27 |
25 |
30 |
8 |
29 |
|
|
S. epidermidis |
15 |
19 |
28 |
28 |
22 |
9 |
17 |
||
|
Surface
Water |
N1 |
S. aureus |
23 |
15 |
26 |
26 |
28 |
23 |
32 |
|
S. epidermidis |
23 |
18 |
29 |
27 |
28 |
12 |
21 |
||
|
N2 |
S. aureus |
23 |
26 |
24 |
22 |
26 |
22 |
31 |
|
|
S. epidermidis |
16 |
19 |
26 |
13 |
21 |
30 |
27 |
||
|
Coastal
Water |
S1 |
S. aureus |
20 |
13 |
27 |
30 |
25 |
32 |
25 |
|
S. epidermidis |
21 |
18 |
21 |
21 |
24 |
16 |
23 |
||
|
S2 |
S. aureus |
28 |
20 |
33 |
32 |
26 |
32 |
33 |
|
|
S. epidermidis |
21 |
15 |
25 |
23 |
23 |
11 |
15 |
||
|
Legend:
FOX-30: Cefoxitin; P-10: Penicillin G, CIP-5: Ciprofloxacin, NOR-10:
Norfloxacin, CN-10: Gentamicin, SXT-25: Trimethoprim-sulfamethoxazole, C-30:
Chloramphenicol. |
|||||||||
3.2. Behavior of Staphylococcus to antibiotics
3.2.1. Sensitivity/resistance of isolated groundwater species
The results of antibiotic susceptibility tests showed that the different staphylococcal strains isolated varied resistances depending on the study site and the antibiotic used. In groundwater, all strains of S. aureus and S. epidermidis isolated from boreholes (B1 and B2) were resistant (100%) to cefoxitin and penicillin G (beta-lactam family). The S. aureus strains showed intermediate susceptibility to ciprofloxacin (100%) (quinolone family) and trimethoprim-sulfamethoxazole (diaminopyrimidine/sulfonamide family) (Figs. 3 B1 and 2B2). In contrast, the S. epidermidis strains isolated from boreholes B1 and B2 were also resistant to ciprofloxacin (100%) and trimethoprim-sulfamethoxazole (100%), respectively. However, the strain isolated from borehole B1 showed intermediate susceptibility to trimethoprim-sulfamethoxazole. With the exception of ciprofloxacin, to which the S. epidermidis strain from borehole B2 was sensitive, all strains were sensitive (100%) to norfloxacin, gentamicin, and chloramphenicol. In the well water (Figs. 3W1 and 3W2), all isolated Staphylococcus strains were resistant (100%) in both wells to cefoxitin and penicillin G, and only to ciprofloxacin (100%) in well W1 for both staphylococcal species; to trimethoprim-sulfamethoxazole (100%) and chloramphenicol (100%) in well W2 for the S. aureus and S. epidermidis strains, respectively. In well W1, S. aureus exhibited intermediate susceptibility (100%) to trimethoprim-sulfamethoxazole; however, this intermediate susceptibility was observed in well W2 with ciprofloxacin for all isolated staphylococcal strains. Furthermore, all staphylococcal strains in both sampling points were susceptible to norfloxacin, gentamicin, and only to chloramphenicol for the S. aureus strains in well W1 and for the S. epidermidis strains in well W2. Thus, it can be observed that the S. aureus and S. epidermidis strains isolated from all the groundwater samples are methicillin-resistant due to their resistance to cefoxitin and exhibit roughly the same antibiotic susceptibility profile.

Figure 3. Distribution of percentages of sensitivity/resistance to tested antibiotics on Staphylococcus aureus and Staphylococcus epidermidis strains from underground waters (FOX-30: Cefoxitin; P-10: penicillin G; CIP-5: ciprofloxacin; NOR-10: norfloxacin; CN-10: gentamicin; SXT-25: trimethoprim-sulfamethoxazole; C-30: chloramphenicol).
3.2.2. Sensitivity/resistance of isolated surface water species
In the surface waters (Figs. 4N1 and 4N2), S. aureus was resistant to penicillin G (100%) only at sampling point N1. No antibiotic resistance was observed at point N2 for the tested S. aureus strain. S. epidermidis was resistant to cefoxitin and penicillin G (100%) at both sampling points. S. epidermidis also showed resistance to trimethoprim-sulfamethoxazole at point N1, and norfloxacin and gentamicin at point N2 (100%). Furthermore, both staphylococcal species showed intermediate susceptibility to ciprofloxacin (100%) at all sampling points in the watercourse, whereas, S. epidermidis was sensitive to chloramphenicol (100%). In S. aureus, sensitivity to cefoxitin (100%), trimethoprim-sulfamethoxazole, norfloxacin, gentamicin, and chloramphenicol was observed at point N1. In contrast, in S. epidermidis, sensitivity at point N1 was primarily observed for norfloxacin and gentamicin, and for trimethoprim-sulfamethoxazole at point N2. Thus, none of the S. aureus strains isolated from these waters were methicillin-resistant. However, the S. epidermidis strains isolated from these waters did not exhibit the same antibiotic susceptibility profile but were methicillin-resistant.

Figure 4. Distribution of percentages of sensitivity/resistance to tested antibiotics on Staphylococcus aureus and Staphylococcus epidermidis strains from surface waters (FOX-30: Cefoxitin; P-10: penicillin G; CIP-5: ciprofloxacin; NOR-10: norfloxacin; CN-10: gentamicin; SXT-25: trimethoprim-sulfamethoxazole; C-30: chloramphenicol).
3.2.3. Sensitivity/resistance of isolated coastal water species
In the coastal waters (Figs. 5S1 and 5S2), all S. aureus strains isolated from the different sampling points were resistant to penicillin G (100%). Only the S. aureus strain isolated from point S1 was resistant to cefoxitin (100%). S. epidermidis showed 100% resistance to both cefoxitin and penicillin G at all the sampling points. S. epidermidis also exhibited 100% resistance to ciprofloxacin at point S1, and to trimethoprim-sulfamethoxazole and chloramphenicol at point S2. S. aureus showed intermediate susceptibility (100%) to ciprofloxacin at point S1 and trimethoprim-sulfamethoxazole (100%) at point S2. In contrast, S. epidermidis showed intermediate susceptibility (100%) to trimethoprim-sulfamethoxazole at point S1 and intermediate susceptibility to ciprofloxacin at point S2. All S. aureus strains were susceptible (100%) to trimethoprim-sulfamethoxazole, norfloxacin, gentamicin, and chloramphenicol. In addition to susceptibility to these antibiotics, the S. aureus strain isolated at point S2 was also susceptible to cefoxitin. However, the S. aureus strain isolated at point S1 was methicillin-resistant, whereas the strain isolated at point S2, was resistant to penicillin. It is also noted that the strains of S. epidermidis isolated from these waters do not have the same antibiotic susceptibility profile, but are still methicillin-resistant.

Figure 5. Distribution of percentages of sensitivity/resistance to tested antibiotics on Staphylococcus aureus and Staphylococcus epidermidis strains from coastal waters (FOX-30: Cefoxitin; P-10: penicillin G; CIP-5: ciprofloxacin; NOR-10: norfloxacin; CN-10: gentamicin; SXT-25: trimethoprim-sulfamethoxazole; C-30: chloramphenicol).
3.3. Impact of proximity to the sea on the susceptibility of microorganisms in coastal waters
3.3.1. Correlations between the parameters studied
Correlations between proximity to the sea and the inhibition diameter of different antibiotics tested on isolated Staphylococcus strains were performed using Spearman's rank correlation test ("r"). Across all stations considered in underground, surface, and coastal waters, a significant and positive correlation (r= 0.427) was observed between proximity to the sea and the inhibition diameter of gentamicin (p<0.05) (Table 3).
Table 3. Correlations between proximity to the sea and the inhibition diameter of the different antibiotics used.
Antibiotics | FOX 30 | P 10 | CIP 5 | NOR 10 | CN 30 | SXT 25 | C30 |
Sea distance | -0,077 | -0,399 | -0,192 | -0,049 | 0,427* | -0,312 | 0,075 |
*: P<0.05 **: P<0.01 | |||||||
4. Discussion
4.1. Susceptibility according to antibiotic families of the isolated germs
The different species of Staphylococcus aureus and Staphylococcus epidermidis showed a high percentage of resistance to the tested antibiotics from different families, depending on the study sites. In the β-lactam family, the percentage of antibiotic resistance in different species of Staphylococcus aureus was higher than 50% on average across all biotope types, with total resistance to methicillin in underground waters. Thus, higher resistance was noted in underground waters compared to coastal and surface waters; the most sensitive species were those from surface waters, particularly with cefoxitin. All the different species of Staphylococcus epidermidis, showed resistance to all tested β-lactamines (100%), regardless of the biotope, and thus all were resistant to methicillin. This could be explained, in part, by the fact that all Staphylococcus epidermidis species produce β-lactamases and are naturally resistant to methicillin. This resistance could be linked to the overexpression of an additional penicillin-binding protein (PBP) with low affinity for beta-lactams, PBP2a. The PBP2a production leads to resistance to all beta-lactams (except ceftobiprole and ceftaroline, which are active against methicillin-resistant staphylococci) [19, 17]. Indeed, this PBP is encoded by the mecA gene, whose complex, called SSCmec (Staphylococcus chromosomal cassette), forms an island of resistance to beta-lactams [20]. On the other hand, this resistance could also be due to the overproduction of plasmid-mediated β-lactamases by S. epidermidis, which are responsible for methicillin hydrolysis [21].
In the quinolone and aminoglycoside families, the antibiotic susceptibility rates for different species of Staphylococcus aureus and Staphylococcus epidermidis were all greater than 50% on average, regardless of the biotope. According to the recommendations of the Antibiogram Committee of the French Society for Microbiology [17], all staphylococcal strains susceptible to norfloxacin are susceptible to fluoroquinolones. This contradicts the findings of Mouhoub [22], who noted that methicillin resistance in staphylococci is accompanied by resistance to all β-lactams as well as other antibiotics, including aminoglycosides and fluoroquinolones. Similarly, Deplano [23], also noted that methicillin resistance in Staphylococcus epidermidis confers resistance to fluoroquinolones. The gentamicin resistance of Staphylococcus epidermidis observed in surface water body N2 (located near a health center) could be due to a one-time contamination by a hospital-acquired strain resistant to aminoglycosides through the secretion of the constitutive enzyme AAC (6')-APH (2'') (aminoglycoside N-acetyl transferase), which confers resistance to all aminoglycosides, including gentamicin [19]. The likely hospital origin of this S. epidermidis strain could also explain this resistance, since, according to Deplano [23], the gentamicin resistance rate for hospital-acquired S. epidermidis strains varies from 25% to over 50%.
In the phenicol family, the percentage of antibiotic sensitivity in the different species of Staphylococcus aureus was 100% in all biotope types. In contrast, the average resistance to Staphylococcus epidermidis was 25%. This indicates low-level resistance to oxazolidinones, according to the Antibiogram Committee of the French Society of Microbiology [17]. This contradicts the findings of Mouhoub [22], who noted that methicillin-resistant strains are those that exhibit multidrug resistance to most antibiotics active against staphylococci, including chloramphenicol, erythromycin, and β-lactamines, among others. In the diaminopyrimidine/sulfamide family, the percentages of antibiotic susceptibility in the different species of Staphylococcus aureus were all greater than 50% on average, regardless of the biotope. In contrast, the average resistance to Staphylococcus epidermidis was 50%. This deviates from the work of [24], who noted that a combination of sulfamethoxazole and trimethoprim was ineffective against coagulase-negative staphylococci, such as Staphylococcus epidermidis and active against methicillin-sensitive staphylococcal strains [25].
4.2. Relationships between the evaluated parameters
Spearman's rank correlation test ("r") showed significant relationships between the proximity of the different sampling points to the sea and certain antibiotics, demonstrating the impact of proximity to the sea on the mechanism of action of antibiotics. The distance between the different sampling points and the sea was significantly (P < 0.05) and positively correlated with gentamicin inhibition diameters. This would mean that most of the tested staphylococci were community-acquired; despite their methicillin resistance, they would not possess aminoglycoside resistance genes and therefore lack the constitutive enzyme AAC (6')-APH (2'') (aminoglycoside N-acetyl transferase), which confers aminoglycoside resistance, as staphylococci are naturally susceptible to it. Proximity to the sea could facilitate the transfer of pollutants (via urban wastewater discharges, ships, and hospitals, etc.) to the different sampling points, which would impact the abiotic parameters of the waters studied and, consequently, the inhibitory effect of gentamicin on the different staphylococcal species. Therefore, these coastal environmental factors would therefore limit the spread of staphylococci carrying the aminoglycoside resistance gene. This contradicts with the findings of Fortun-Lamothe [26], who noted that environmental factors play a role in the emergence of antibiotic resistance in certain bacteria.
5. Conclusions
This study aimed to determine the susceptibility of two Staphylococcus species and the potential impact of proximity/contact with seawater on the susceptibility of these two Staphylococcus species to antibiograms. These species were isolated from coastal groundwater and surface water in Kribi, Cameroon. Bacteriological analyses revealed the presence of a bacterial microflora consisting of Staphylococcus aureus and S. epidermidis. Across the aquatic systems, the majority of isolated S. aureus and S. epidermidis strains were found to be methicillin-resistant due to their resistance to cefoxitin. The different Staphylococcus species isolated from groundwater exhibited complete resistance to antibiotics, such as beta-lactams. Antibiotic resistance in different families was higher in staphylococcal species isolated from groundwater compared to those isolated from surface and coastal waters. Proximity to or contact with seawater at the sampling points appeared to have a greater influence on the gentamicin susceptibility of Staphylococcus bacteria, as the tested strains were likely community-acquired and lacked the enzymes responsible for aminoglycoside resistance.
Disclaimer (artificial intelligence)
Author(s) hereby state that no generative AI tools such as Large Language Models (ChatGPT, Copilot, etc.) and text-to-image generators were utilized in the preparation or editing of this manuscript.
Authors’ contributions
Conceptualization, P.L., O.V.N.; methodology, P.L., O.V.N., M.N.; software, P.L., O.V.N., M.N.; validation O.V.N., G.B., P.A.N., T.S.N., M.N.; formal analysis, P.L., O.V.N., M.N.; investigation, P.L., T.I.B.N., S.D.B., Y.P.Y., C.S.M.; resources, P.L., O.V.N. ; data curation, P.L., O.V.N., T.I.B.N., S.D.B., Y.P.Y., C.S.M.; writing– original draft and preparation, P.L., O.V.N., M.N.; writing – review & editing, P.L., O.V.N., M.N., visualization, O.V.N., G.B., P.A.N., T.S.N., M.N.; supervision, M.N.; project administration, O.V.N., G.B., P.A.N., T.S.N., M.N.; funding acquisition, P.L., O.V.N., T.I.B.N., S.D.B., Y.P.Y., C.S.M., G.B., P.A.N., T.S.N., M.N.
Acknowledgements
Our thanks to the Hydrobiology and Environment Laboratory of the University of Yaoundé 1.
Funding
This research received no specific grant from any funding agency.
Availability of data and materials
All relevant data are within the paper. Additional data will be made and available on request according to the journal policy.
Conflicts of interest
The authors declare no potential conflict of interest regarding the publication of this work.
References
This work is licensed under the
Creative Commons Attribution
4.0
License (CC BY-NC 4.0).
Abstract
Kribi is one of Cameroon's most popular seaside resorts. Its beaches are generally fed by rivers and are sites of numerous human activities. Aquatic environments are increasingly subject to multiple forms of human-induced contamination. The pathogenic microorganisms present can cause illness and economic losses in activities that depend primarily on seawater. This study aimed to determine the susceptibility of two species of the genus Staphylococcus and the potential impact of proximity/contact with seawater on the antibiotic susceptibility of these two Staphylococcus species isolated from underground and surface coastal waters at Kribi (Cameroon). The bacteria were isolated using the surface spread technique on Petri dishes with Chapman mannitol agar (HIMEDIA, ref. MH118-500G). The isolated bacteria were identified using standard methods. The susceptibility of the bacteria to antibiotics was verified using the disk diffusion method on Müller-Hinton agar, following the recommendations of the Antibiogram Committee of the French Society of Microbiology. Bacteriological analyses revealed that these waters host a bacterial microflora composed of Staphylococcus aureus and Staphylococcus epidermidis. Antibiotic susceptibility tests showed that the different bacterial species exhibited resistance to methicillin (100%) (due to their resistance to cefoxitin) and penicillin G, which belongs to the β-lactam family. Within the quinolone, aminoglycoside, phenicol, and diaminopyrimidine/sulfamide families, the percentages of antibiotic susceptibility for the different species of Staphylococcus aureus and Staphylococcus epidermidis were all greater than 50% on average, regardless of the biotope. The percentage of staphylococcal resistance in underground and coastal waters was higher (78.57%), compared to surface waters (71.43%). The susceptibility to gentamicin in the different isolated staphylococcal species appeared to be favored by their proximity or contact with seawater, thus showing the influence of abiotic factors on the antibiotic susceptibility of bacterial species.
Abstract Keywords
Surface water, underground water, coastal water, Staphylococcus, antibiotic.
This work is licensed under the
Creative Commons Attribution
4.0
License (CC BY-NC 4.0).
Editor-in-Chief

This work is licensed under the
Creative Commons Attribution 4.0
License.(CC BY-NC 4.0).